
|
This is the lab page of David T Blake, PhD. The lab investigates questions concerning brain electrophysiology and behavior. The most recent focus has been on basal forebrain deep brain stimulation, its physiological and behavioral effects, and the potential to use such deep brain stimulation to treat neurological disorders. In multiple studies in Rhesus monkeys, aged Rhesus monkeys, mice, and humans we routinely measure cognitive changes from deep brain stimulation equivalent to reversing the age related declines in cognition over the last third of the lifespan. Honestly, I think it is going to cure Alzheimer's, and by that I mean its application in patients will prevent cognitive decline from altering their activities of daily living. Human trials of our methods are ongoing in some centers. If you would like to apply our methods to your human trials, contact me. |
Main questions for the lab |
| Weeks of application of deep brain stimulation can dramatically improve executive function even in fully functioning young adults. Can it do the same in Alzheimer's patients? |
| What are the cell biological pathways involved in instantiating those learning rules? |
| The release of acetylcholine and noradrenaline by deep brain stimulation of the basal forebrain or locus coeruleus cause alter neurotrophic tone, neural tone, and blood vessel diameter. What are the causal and dependent relationships between local neural activity, blood vessel tone, and neuromodulation? |
|
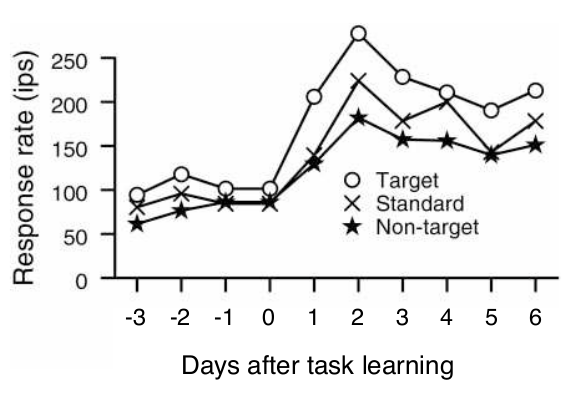
In 1996, I went to the lab of Michael Merzenich. There, with R. Christopher deCharms, we succeeded in developing one of the earliest high yield sensory cortex implants. The following experiments, done also with fabrizio strata, Anne Churchland, and Richard Kempter, were among the first to resample, with high spatial resolution, the same cortical locations throughout a period of learning. The discoveries that followed were unexpected. A robust nonselective response enhancement occurred within 24 hours of the time that an animal learned to associate a new sensory stimulus with reward. Over longer periods of time, the representations of stimuli associated with omission of reward, or task distractors, became selectively suppressed. Surprisingly, any tendency for the representations of targets to be enhanced, relative to the representations of control stimuli, were weak or absent. The following figure, taken from Blake et al 2002, shows the rapidity and strength of the nonselective response enhancement. Cortical responses double in 24 hours after learning. |

|
Current work occurs in the mouse lab, led by Khadijah Shanazz,
PhD.
She uses the GACh3.0 sensor in cerebral cortex, and also places a
stimulating electrodes in the basal forebrain. She can stimulate
and image the cholinergic response to stimulation each 0.6
seconds.
One paper is on BioRXiV, and she has recently added two
things to her studies. She measures the vascular response to
stimulation at the same time as the cholinergic response, and she
transitioned to working with awake mice. We've received a lot of
help from the labs of Sergei Kirov and Phil O'Herron in getting
things working, and they are supportive co-authors. Dr. Shanazz can be emailed as kshanazz AT augusta |

|
The work in the monkey lab is led by graduate student Kendyl
Pennington. She has a paper
on BioRXiV also, and hers details the
changes in senescent macaques after deep brain stimulation of the
nucleus basalis of Meynert is applied. Her upcoming project
samples cerebrospinal fluid before, during, and after deep brain
stimulation. She is looking at changes in the proteome of the
fluid as indications of processes occuring in the chemical
communication between neurons and other cells in the neuropil. We
are particularly interested if there are clues to the causes of
Alzheimer's dementia that are reflected by the proteome of the
CSF fluid. Stay tuned for answers! Kendyl can be emailed as kpennington AT augusta |